Measuring the weak light re-emitted by cells and organisms may tell us a lot about them. Dr. Mae-Wan Ho reports
I first became aware of some unusual research at a conference organised by Clive Kilmister, Emeritus Professor of Mathematics, King's College, London, in 1985, on "Disequilibrium and Self-organisation". There, I met, among others, German physicist Fritz Popp, whose talk meant almost nothing to me then, except for the claim that organisms are "coherent", and that the proof of coherence was in the characteristics of the extremely weak after-glow that organisms emit immediately after briefly stimulated with light.
That meeting changed my whole field of research, thanks to collaborative work with Fritz Popp, who taught me a lot about quantum physics, and later, with Franco Musumeci's team in Catania University, Sicily, which continues my incredible voyage of discovery.
I later learned that most materials glow in the dark after they have been exposed to light, but the after-glow emitted by all living cells and organisms is different. It is also different from the much more intense fluorescence exhibited by certain chemical compounds, or the strong flashes of light emitted by fireflies due to special biochemical reactions that generate light.
Living cells and organisms also emit extremely low levels of light spontaneously, as Alexander Gurvich discovered in Russia in 1923. He thought this light was involved in intercommunication between cells.
Fifty years later, Fritz-Albert Popp built the first photon detector sensitive enough to study these "biophotons", as he calls them. Popp, too, believes cells and organisms use biophotons to intercommunicate. Moreover, he thinks that both the spontaneously emitted biophotons as well as the after-glow stimulated by external light are coherent, and come from a coherent "light-field" in the cells and organisms. In other words, biophotons represent an extremely weak as well as a most unusual laser light emitted by the living system; a laser that covers a broad range of frequencies, from the ultra-violet to the infrared, and probably beyond, into the microwave and radio-frequency range.
Although many laboratories have been able to detect biophotons, the coherence of biophotons is much disputed. Many scientists believe they are no more than the result of "imperfections" or "mistakes" in the biochemical reactions taking place in the body. But decades of empirical research by Popp and others have shown without doubt that all cells and organisms emit biophotons, the characteristics of which are intimately dependent on their physiological state.
There is also a lot of other evidence indicating that organisms are highly coherent, if not quantum coherent (see The Rainbow and the Worm, The Physics of Organisms – thereafter abbreviated to "The Rainbow Worm" - now available from ISIS' online store).
Some of the most revealing findings on biophotons were made in connection with food and other agricultural products. Popp's laboratory pioneered food quality research with support coming from some of the biggest food companies. He and his coworkers found it possible to distinguish organic tomatoes from conventionally grown tomatoes from a supermarket. Similarly, free-range eggs could be distinguished from battery-hen eggs, and the germination rate of barley seeds could be predicted from their after-glow.
Popp's work has inspired many other laboratories around the world. Some have devoted major efforts to assessing food quality. It is not hard to understand why a simple, non-destructive method such as biophoton emission is needed.
For example, cherry tomatoes (Lycopersicon esculentum var. cerasiforme) are harvested at various stages to ripen on storage. The storage-ripened tomatoes looked the same in colour, size and degree of firmness, but human tasters are able to distinguish those picked earlier as less sweet and less tasty, and having more "off-flavour" than those picked later. And this could be confirmed by chemical methods to assess sugar and solid contents. The drawback is that the chemical tests destroy the tomatoes and many of them are costly and time-consuming to carry out.
By measuring the after-glow, the research team led by Franco Musumeci in Catania University was able to distinguish the earlier picked tomatoes without any difficulty.
The after-glow or "delayed luminescence" (DL) is measured within a hundred milliseconds (or earlier) after the brief pulse of stimulating light is off. DL typically starts at a high level, and decays 'hyperbolically' (as a function of time) back to the background in seconds, or sometimes minutes.
Musumeci and his colleagues found, first of all, that the intensity of the after-glow decreased as the tomatoes matured, and was closely correlated with the decrease in the rate of respiration as well as the increase in the redness of the tomatoes. In other words, the greener, and less mature the tomatoes, the higher the DL. The DL of all samples dropped during the storage period.
By the end of the storage period of ten days, all the tomatoes were the same indistinguishable shade of red. But striking differences remained in the intensity DL, which decreased with the maturity of the fruit at harvesting and the increase in sugar content (or "Solids", see Fig. 1).
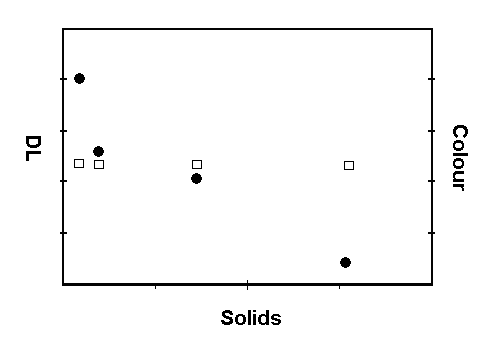
Figure 1. Delayed luminescence and colour as a
function of solids in cherry tomatoes.
Seed quality is as important as food quality, if not more so, as the profitability of agriculture depends on it. The farmer wants a seed lot that give a high rate of germination quickly and the resulting seedlings to develop into vigorous plants. Stress can affect seed viability and vigour during all stages of production, harvesting drying, storage, packaging and transport.
In a study carried out in 1994, Musumeci's group had established that soybean seeds heat stressed for varying periods of time had decreased growth rates proportional to the length of heat stress, while the intensity of DL increased proportionately. There were also significant disturbances to the decay times of the DL, tending to shorten it.
Research carried out by others have shown that seeds improve and recover vigour after 'priming', or being soaked in osmotically active agents such as polyethylene glycol (PEG). Many molecular and physiological processes are correlated with the loss of seed vigour, among which, the accumulation of gene and chromosome mutations, loss of integrity of ribosomal RNA, decrease in membrane phospholipid content and increase in fatty acids.
The improvement in vigour following priming was correlated with completion of DNA repair during priming and a more favourable metabolic balance of the primed seeds at the start of germination in water.
Studies on capsicum pepper seeds carried out by the Plant Breeding and Seed Production group in the University of Torino, Italy, showed that cells of the embryo in the dried seeds arrest the cell cycle at the 'G1 phase', before DNA is synthesized in the nucleus. Priming in PEG solutions induced DNA synthesis in the embryo root tips. Within each seed lot, a significant direct correlation was seen between the frequency of priming-induced nuclear replication and the improvement in seed vigour, as measured by the reduction in the mean time to germination. But, the amount of priming-induced nuclear replication was also correlated with the degree of seed deterioration, so nuclear replication by itself may not be a reliable guide of improvement or seed vigour.
Musumeci's group teamed up with researchers in Torino to investigate whether delayed luminescence could (literally) throw further light on the issue.
They found that keeping the seeds at 45C for 4 and 6 days led to a progressive increase in intensity of DL compared with controls, which was significant after 6 days. No significant changes in germination rate was observed, while the mean germination time increased from 5 days in controls to 7.8 days after 6 days at 45C.
Priming control seeds for 6 and 12 days had no significant effect on the germination rate, but significantly shortened the mean germination time to 3.5 and 2.9 days respectively. The percent of nuclei entering G2 phase (making DNA) increased from 0 to 8.5% and 20% respectively. This was accompanied by significant decreases in the intensity of DL.
The seeds kept at 45C for 4 days responded to priming for 6 days by a significant shortening of the mean germination time from 5.5 days to 3.3 days. No nuclei had entered the G2 phase, but the DL had decreased significantly, indicating that the improvements in priming were independent of DNA synthesis. After priming for 12 days, germination rate
decreased from 92% in the unprimed seeds to 83%, 13.6% of the nuclei entered G2 phase and there was a further reduction in the DL.
Seeds kept at 45C for 6 days and primed for 6 days showed a slight shortening of mean germination time and a small reduction in the intensity of DL, again without any nuclei entering G2 phase. After priming for 12 days, the mean germination time had shortened from 7.8 days to 3.4 days, the germination rate had also gone down from 92 to 85.5%, 10.1% of the nuclei had entered G2 phase and DL had decreased sharply.
Further analysis of the data showed that the intensity of DL was highly correlated with mean germination time, that is, the longer the mean germination time, the higher the DL.
So DL appears to be a better predictor of some aspects of seed vigour than nuclear replication.
The orthodox scientific community has difficulty understanding these results. They are used to the idea that light is emitted by specific light-emitting molecules (or chromophores). But that is immediately contradicted by the fact that biophotons consist of light of a wide, continuous range of frequencies, rather than a single or a few frequencies as would be the case if special light-emitting molecules were involved.
All the findings indicate that biophotons come from the entire cell or organism, which is behaving rather like a special 'solid state' device with energy stored throughout the system, as I have suggested in The Rainbow Worm.
The external light goes to excite the system as a whole. The excited energy is distributed throughout the system and eventually part of it is re-emitted as light over a wide band of frequencies, reflecting the complex excitation state of the whole.
Of course, the analogy with a solid-state device - which originated with solid-state physicist Herbert Fröhlich - is very crude, like the analogy of biophotons with laser light. First of all, the organism, and even the single cell, has a complex, nested organisation that's unrivalled in any artificial solid-state device. Cells have their own skeleton, compartments, and tiny organs studded with molecular machines turning autonomously, transforming energy. More importantly, organisms are definitely not solids but liquid crystalline, consisting of 70% by weight of water, which is increasingly recognized by scientists in the mainstream as the most important constituent of living systems. (An entire Royal Society discussion meeting was recently devoted to the question: is life possible without water?) The large amounts of water associated with living organisms offer the flexibility that practically all proteins, DNA, RNA and other macromolecules need in order to work at all, or to work to the high efficiency required in the organisms.
Nevertheless, the analogy is useful, as solid-state systems do exhibit DL similar (though not exactly the same) to that emitted by organisms and cells. Musumeci's group discovered that the intensity of DL is a function of the size of the grains, and therefore, of the domains in which the fine-order physical structure can sustain a band of excited electronic levels that gives rise to DL. When the grains are reduced to powder, DL disappeared. The standard explanation for DL in solid-state systems is that the excited electrons move from their fixed orbits around the nucleus of the atoms, and eventually fall back to the 'ground state'. In the process, some of the energy of excitation is re-emitted as delayed luminescence, while the rest is dissipated as heat.
In collaboration with Musumeci's team, we have shown that DL disappears, or is reduced to a very low level when the organisation of the cell is disrupted by homogenisation, or by immersing in an ionic medium that irreversibly fragments the cytoskeleton (skeleton of the cell made of special fibre-forming proteins).
Similarly, using slices of the isolated beef Achilles tendon, we were able to show that DL is closely dependent on the structure of the collagen fibrils and the associated biological water. As the water content decreases, drastic changes take place both in the intensity of the DL and in its rate of decay (slope of the hyperbolic decay curve).
In the cell, as in the solid-state system, excitation energy can make electrons or even protons (positively charged hydrogen nucleus) move through the system, and there are many other ways in which the excitation energy can be stored transiently, before it is dissipated, as heat or light: vibrations of chemical bonds, large fluctuations of protein and other macromolecules and electrical currents through the cells and organisms, to mention but a few. Intuitively, one can see that the more the cell or organism has the capacity to store the energy, the less will be re-emitted, and also the more long-lasting is the DL (the slope of the hyperbolic decay curve is less steep). That may be why there is an inverse relationship between the vigour of seeds and intensity of DL. But that is no more than a hypothesis at the moment.
It is clear that a lot more research on non-destructive physical methods is needed. An imaging technique, Symchromics©, invented in my laboratory allows us to see all living organisms in brilliant colours, and offers the possibility of measuring the coherence of the organism and cells directly.
The colours depend on the motions of the molecules in tissues and cells being highly coherent. Because light vibrates much faster than the coherent motion of the molecules, living organisms look as if they are made of statically aligned liquid crystals, thereby generating the same kind of 'interference colours' that are produced by rock crystals.
Indeed, the intensity of the colours is directly dependent on the coherence of the molecular motions. Significantly, the most active parts of the organism are invariably the brightest parts; which suggests that coherence is a function of the energetic status or vitality of the organism. The colours fade and disappear as the organism dies, which is when random molecular motions take over.
What does coherence amount to in the organism in terms of energy storage and mobilisation? It amounts to energy being mobilised and distributed throughout the system most rapidly and efficiently, the energy effectively remaining stored as it is mobilised (see "Why are organisms so complex?" this issue). When such a system is perturbed with an external light pulse, its degree of coherence is bound to affect how the energy in the light pulse excites the system, and how that energy is re-emitted as after-glow.
It would be simple and revealing to correlate DL measurements with measurements on coherence using Symchromics©. We fully intend to do that when we can get the funding required.
Article first published 05/01/04
Comments are now closed for this article