Low chronic exposure to the soluble insecticide is lethal to fresh water shrimps Dr Eva Sirinathsinghji
A new study shows neonicotinoid insecticides are lethal to freshwater invertebrates following low but constant exposure. This group of insecticides are in the headlines again following a partial EU–wide ban agreed in April 2013, as evidence of their toxicity to bees became indisputable. Thirty studies have now linked neonicotinoid insecticides to the global demise of bees and is a likely cause of the Colony Collapse Disorder as ISIS was among the first to suggest (see [1] Requiem for the Honeybee, SiS 34, [2] Emergency Pesticide Ban for Saving the Honeybee, SiS 39).
New evidence has now emerged of their toxicity to another species, the arthropod Gammarus pulex, a freshwater shrimp; a predictable effect of a highly water-soluble chemical present in surface waters.
The study, conducted by Dr Roman Ashauer and colleagues from Eawag (the Swiss Federal Institute of Aquatic Science and Technology, Switzerland) looked at how different exposures of Imidacloprid, made by Bayer, would impact feeding activity, lipid content, immobility and survival of the fresh water shrimp [3]. They looked at both pulsed exposure - two high concentration pulses of 90 µg/L with either short or long intervals between pulses - aimed at mimicking natural fluctuations of water contamination, as well as constant low level exposures of the same overall amount as used in the pulse experiments, at 15 µg/L. The shrimp, collected from a stream in Switzerland, were exposed for either 14 or 21 days, revealing effects not seen in routine toxicology tests that do not include long-term exposure.
The results showed significant mortality with constant low-level exposure: 92 and 95 % of insects died in the 14 and 21 day experiments respectively, the remaining 8 and 5 % of living animals were immobilised by the insecticide (see Figure 1). There was a reduction in the proportion of mobile animals over the entire course of the experiment compared with controls; the data was plotted against the predicted outcome from Toxicokinetic-Toxicodynamic (TK-TD) models (see below).
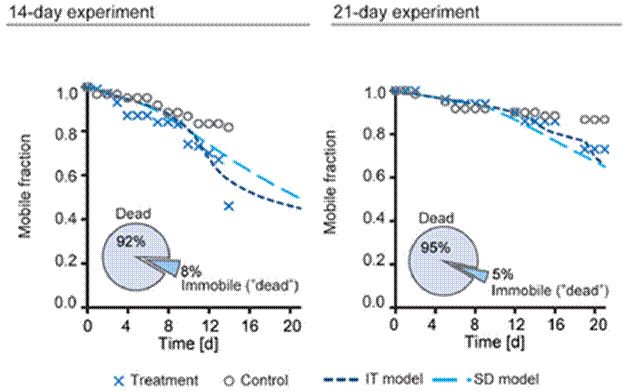
Figure 1 Survival of G. purex after 14 (left) and 21 day (right) constant low level exposure to imadocloprid; survival data plotted alongside predicted survival outcomes from both individual tolerance (IT) and stochastic death (ST) models of survival; percentage of dead or immobile animals presented in the pie charts
Feeding behaviour, as measured by the mass of leaf material eaten was significantly affected, consistent with reduced lipid content in the animals. Figure 2 shows that the untreated animals had a lipid content of 1.4 % compared to exposed animals at 0.7 %, consistent with a significant reduction in feeding. Significant effects are more pronounced after 14 day experiment due to possible variability in background mortality, lipid reserves or fitness levels at different times of the year, the 14 and 21 day experiments being conducted in November and February respectively.
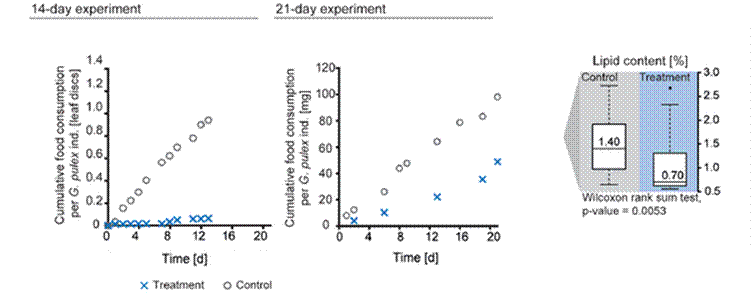
Figure 2 Food consumption of exposed and unexposed G. purex after 14 (left) and 21 day (middle) constant low-level exposure to imadocloprid; reduced lipid content measured after 21 days (right) is consistent with reduced feeding
The decreased feeding rate implies that starvation is at least partially responsible for imidacloprid lethality.
High-level exposure immobilised the animals, but they recovered fast upon transfer to clean water after the pulses, and therefore no significant effect on feeding or lipid content was observed.
The different results obtained through continuous low-level or pulsed high-level exposure reflect the mechanism of neonicotinoids, a known neurotoxin. They bind to nicotinic acetylcholine receptors to interfere with neuronal signalling, particularly to muscle, resulting in twitching, cramps and muscle weakness at lower levels. This can impair feeding behaviour and make animals more passive, as already found in other species [4-6]. Higher level exposure on the other hand, is thought to cause paralysis and death via overstimulation and blockage of the receptors. The differing mechanisms of low and high level exposure may partially explain the divergent results with pulsed experiments, where the animals were mostly immediately paralyzed or killed (less than with constant low exposure) but showed no reduction in feeding behaviour or lipid content if transferred back to clean water where they recovered after 2 days.
Survival data gathered in the initial experiments were applied to Toxicokinetic-Toxicodynamic (TK-TD) models, an integrating concept for the quantification of toxic effects. They simulated the processes consistent with the data, taking into account internal chemical concentrations, internal damage, rate of internal chemical elimination and concentration of exposure among other things. The survival data of both pulsed and constant exposures fitted best with an individual tolerance model (IT), which assumes that individual organisms react differently to any given chemical, in this case it could be down to individual lipid or other energy reserves that would protect from starvation for example. This simulation approach also suggested as predicted that different mechanisms underlie toxicity for constant low level exposure versus pulsed high level exposure.
Also investigated was whether starvation was the sole factor in the observed lethality in the initial experiments. Starvation experiments were performed without insecticide and this data was compared to the TK-TD IT model calibrated for limited food availability. They found that the starvation model could not accurately predict the results seen with the constant exposure, suggesting that additional factors are involved in killing the animals. As acetylcholine neurotransmission is involved in other key physiological processes such as respiration, the authors suggest that these may also contribute to the toxicity of imadocloprid.
Based on their data, the authors developed a model for simulating survival under constant exposure, called the multiple stress model that combines the effects of starvation and other toxic pathways of the chemical that was able to accurately reproduce their survival data.
This study and the new modelling approach need to be considered by regulators. Current regulatory assessments are inadequate and would have missed the effects observed in this work. Long-term studies such as those performed here are still not required for regulatory approva;.
Article first published 19/06/13
Comments are now closed for this article