Mae-Wan Ho
Bioelectrodynamics Laboratory,
Open University, U.K.
Journal of Consciousness Studies 3, 231-244, 1996
Abstract: According to Bergson (1916), the traditional problem of free will is misconceived and arises from a mismatch between the quality of authentic, subjective experience and its description in language, in particular, the language of the mechanistic science of psychology. Contemporary western scientific concepts of the organism, on the other hand, are leading us beyond conventional thermodynamics as well as quantum theory and offering rigorous insights which reaffirm and extend our intuitive, poetic, and even romantic notions of spontaneity and free will. I shall describe some new views of the organism arising from new findings in biology, in order to show how, in freeing itself from the 'laws' of physics, from mechanical determinism and mechanistic control, the organism becomes a sentient, coherent being that is free, from moment to moment, to explore and create its possible futures.
*Based on a lecture delievered at the 6th Mind & Brain Symposium, The Science of Consciousness - The Nature of Free Will, November 4, 1995, Institute of Psychiatry, London.
Distinguished neurophysiologist Walter Freeman (1995) begins his latest book by declaring brain science 'in crisis': his personal quest to define constant psycho-logical states arising from given stimuli has ended in failure after 33 years. Patterns of brain activity are simply unrepeatable, every perception is influenced by all that has gone before. The impasse, he adds, is conceptual, not experimental or logical. This acknowledged breakdown of mechanical determinism in brain science is really long overdue, but it should not be miscontrued as the triumph of vitalism. As Freeman goes on to show, recent developments in nonlinear mathematics can contribute to some understanding of these non-repeatable brain activities.
The traditional opposition between mechanists and vitalists already began to dissolve at the turn of the present century, when Newtonian physics gave way to quantum theory at the very small scales of elementary particles and to general relativity at the large scales of planetary motion. The static, deterministic universe of absolute space and time is replaced by a multitude of contingent, observer-dependent space-time frames. Instead of mechanical objects with simple locations in space and time, one finds delocalized, mutually entangled quantum entities that carry their histories with them, like evolving organisms. These developments in contemporary western science gave birth to organicist philosophy.
A key figure in organicist philosophy was the French philosopher, Henri Bergson (1916), who showed how Newtonian concepts - which dominate biological sciences then and now - negate psychology's claims to understand our inner experience at the very outset. In particular, he drew attention to the inseparability of space and time, both tied to real processes that have characteristic durations. The other major figure in organicist philosophy was the English mathematician-philosopher, Alfred North Whitehead (1925) who saw physics itself and all of nature, as unintelligible without a thorough-going theory of the organism that participates in knowing.
Organicist philosophy was taken very seriously by a remarkable group of people who formed the multidisciplinary Theoretical Biology Club [1]. Its membership included Joseph Needham, eminent embryologist/biochemist later to be renowned for his work on the history of Chinese science; Dorothy Needham, muscle physiologist and biochemist, geneticist C.H. Waddington, crystallographer J.D. Bernal, mathematician Dorothy Wrinch, philosopher, J.H. Woodger and physicist, Neville Mott. They acknowledged the full complexity of living organization, not as axiomatic, but as something to be explained and understood with the help of philosophy as well as physics, chemistry, biology and mathematics, as those sciences advance, and in the spirit of free enquiry, leaving open whether new concepts or laws may be discovered in the process.
A lot has happened since the project of the Theoretical Biology Club was brought to a premature end when they failed to obtain funding from the Rockefeller Foundation. Organicism has not survived as such, but its invisible ripples have spread and touched the hearts and minds, and the imagination of many who remain drawn to the central enigma that Erwin Schrödinger (1944) later posed: What is Life?
In the intervening years, the transistor radio, the computer and lasers have been invented. Whole new disciplines have been created, nonequilibrium thermodynamics, solid state physics and quantum optics to name but a few. In mathematics, nonlinear dynamics and chaos theory took off in a big way during the 1960s and 70s. Perhaps partly on account of that, many nonlinear physical and physicochemical phenomena are being actively investigated only within the past ten years, as physics become more and more organic in its outlook.
In a way, the whole of science is now tinged with organicist philosophy, as even 'consciousness' and 'free will' are on the scientific agenda. Bergson (1916) has made a persuasive case that the traditional problem of free will is simply misconceived and arises from a mismatch between the quality of authentic, subjective experience and its description in language, in particular, the language of the mechanistic science of psychology. In a recent book, I have shown how contemporary western scientific concepts of the organism are leading us beyond conventional thermodynamics as well as quantum theory (Ho, 1993), and offering rigorous insights which reaffirm and extend our intuitive, poetic, and even romantic notions of spontaneity and free will.
I am making a case for organicist science. It is not yet a conscious movement but a Zeitgeist I personally embrace, so I really mean to persuade you to do likewise by giving it a more tangible shape. The new organicism, like the old, is dedicated to the knowledge of the organic whole, hence, it does not recognize any discipline boundaries. It is to be found between all disciplines. Ultimately, it is an unfragmented knowledge system by which one lives. There is no escape clause allowing one to plead knowledge 'pure' or 'objective', and hence having nothing to do with life. As with the old organicism, the knowing being participates in knowing as much as in living. Participation implies responsibility, which is consistent with the truism that there can be no freedom without responsibility, and conversely, no responsiblity without freedom. There is no placing mind outside nature as Descartes has done, the knowing being is wholeheartedly within nature: heart and mind, intellect and feeling (Ho, 1994a). It is non-dualist and holistic. In all those respects, its affinities are with the participatory knowledge systems of traditional indigenous cultures all over the world.
From a thorough-going organicist perspective, one does not ask, 'What is life?' but, 'What is it to be alive?'. Indeed, the best way to know life is to live it fully. It must be said that we do not yet have a fully fledged organicist science. But I shall describe some new images of the organism, starting from the more familiar and working up, perhaps to the most sublime, from which a picture of the organism as a free, spontaneous being will begin to emerge. I shall show how
the organism succeeds in freeing itself from the 'laws' of physics, from mechanical determinism and mechanistic control, thereby becoming a sentient, coherent being that, from moment to moment, freely explores and creates its possible futures.
I put 'laws' in quotation marks in order to emphasize that they are not laid down once and for all, and especially not to dictate what we can or cannot think. They are tools for helping us think; and most of all, to be transcended if necessary.
Many physicists have marvelled at how organisms seem able to defy the Second Law of Thermodynamics, starting from Lord Kelvin, co-inventor of the Second Law, who nevertheless excluded organisms from its dominion:
'The animal body does not act as a thermodynamic engine...consciousness teaches every individual that they are, to some extent, subject to the direction of his will. It appears therefore that animated creatures have the power of immediately applying to certain moving particles of matter within their bodies, forces by which the motions of these particles are directed to produce derived mechanical effects.' [2]
What impresses Lord Kelvin is how organisms seem to have energy at will, whenever and wherever required, and in a perfectly coordinated way. Another equally puzzling feature is that, contrary to the Second Law, which says all systems should decay into equilibrium and disorder, organisms develop and evolve towards ever increasing organization. Of course, there is no contradiction, as the Second Law applies to isolated systems, whereas organisms are open systems. But how do organisms manage to maintain themselves far away from thermodynamic equilibrium and to produce increasing organization? Schrödinger writes:
'It is by avoiding the rapid decay into the inert state of 'equilibrium' that an organism appears so enigmatic....What an organism feeds upon is negative entropy, or, to put it less paradoxically, the essential thing in metabolism is that the organism succeeds in freeing itself from all the entropy it cannot help producing while alive.' 3
Schrödinger was severely reprimanded, [4] by Linus Pauling and others, for using the term 'negative entropy', for it really does not correspond to any rigorous thermodynamic entity. However, the idea that open systems can 'self-organize' under energy flow became more concrete in the discovery of 'dissipative structures' (Prigogine, 1967). An example is the Bénard convection cells that arise in a pan of water heated uniformly from below. At a critical temperature difference between the top and the bottom, a phase transition occurs: bulk flow begins as the lighter, warm water rises from the bottom and the denser, cool water sinks. The whole pan eventually settles down to a regular honeycomb array of flow cells. Before phase transition, all the molecules move randomly with respect to one another. However, at a critical rate of energy supply, the system self-organizes into global dynamic order in which all the astronomical numbers of molecules are moving in formation as though choreographed to do so.
A still more illuminating physical metaphor for the living system is the laser (Haken, 1977), in which energy is pumped into a cavity containing atoms capable of emitting light. At low levels of pumping, the atoms emit randomly as in an ordinary lamp. As the pumping rate is increased, a threshold is reached when all the atoms oscillate together in phase, and send out a giant light track that is a million times as long as that emitted by individual atoms. Both examples illustrate how energy input or energy pumping and dynamic order are intimately linked.
These and other considerations led me to identify Schrödinger's 'negative entropy' as 'stored mobilizable energy in a space-time structured system' (Ho, 1994b, 1995a). The key to understanding the thermodynamics of living systems turns out not so much to be energy flow but energy storage under energy flow (Fig. 1). Energy flow is of no consequence unless the energy can be trapped and stored within the system where it circulates to do work before dissipating. A reproducing life cycle, i.e., an organism, arises when the loop of circulating energy is closed. At that point, we have a life cycle, within which stored energy is mobilized, remaining largely stored as it is mobilized.
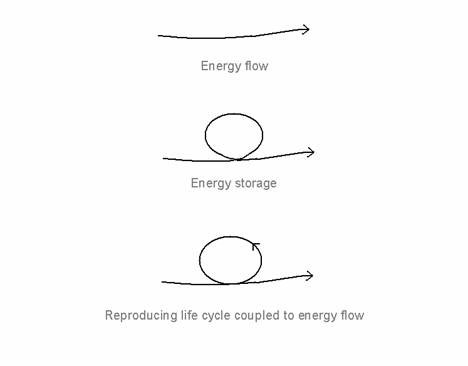
Figure 1. Energy flow, energy storage and the reproducing life-cycle
The life cycle is a highly differentiated space-time structure, the predomi-nant modes of activity are themselves cycles spanning an entire gamut of space-times from the local and fast (or slow) to the global and slow (or fast), all of which are coupled together. These cycles are most familiar to us in the form of biological rhythms extending over 20 orders of magnitude of time, from electrical activities of neurons and other cells to circadian and circa-annual rhythms and beyond. An intuitive picture is given in Figure 2, where coupled cycles of different sizes are fed by the one-way energy flow. This complex, entangled space-time structure is strongly reminiscent of Bergson's 'durations' of organic processes, which necessitates a different way of conceptualizing space-time as heterogeneous, nonlinear, multidimensional and nonlocal (see Ho, 1993). [5]
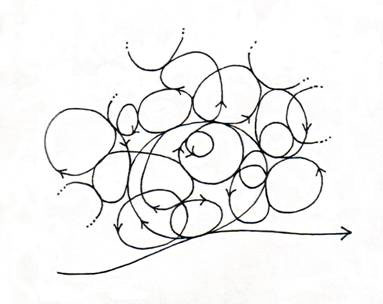
Figure 2. The many-fold cycles of life coupled to energy flow
On account of the complete spectrum of coupled cycles, energy is stored and mobilized over all space-times according to the relaxation times (and volumes) of the processes involved. So, organisms can take advantage of two different ways of mobilizing energy with maximum efficiency - nonequilbrium transfer in which stored energy is transferred before it is thermalized, and quasi-equilibrium transfer, for which the free energy change approaches zero according to conventional thermodynamic considerations (McClare, 1971). Energy input into any mode can be readily delocalized over all modes, and conversely, energy from all modes can become concentrated into any mode. In other words, energy coupling in the living system is symmetrical, which is why we can have energy at will, whenever and wherever required (see Ho, 1993, 1994b, 1995a,b). The organism is, in effect, a closed, self-sufficient energetic domain of cyclic non-dissipative processes coupled to the dissipative processes. In the formalism of conventional thermodynamics, the life cycle can be considered, to first approximation, to consist of all those cyclic processes - for which the net entropy change balances out to zero - coupled to those dissipative processes necessary for keeping it going, for which the net entropy change is greater than zero (see Figure 3). This representation, justified in detail elsewhere (Ho, 1996a), is derived from the thermodynamics of the steady state (see Denbigh, 1951).
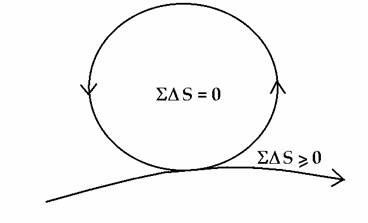
Figure 3. The organism frees itself from the contraints of energy conservation and the second law of thermodynamics
Consequently, the organism has freed itself from the immediate constraints of energy conservation - the First Law - as well as the Second Law of thermodynamics. There is always energy available within the system, which is mobilized at close to maximum efficiency and over all space-time modes. [6]
It was geneticist/embryologist C.H. Waddington (1957) who first introduced nonlinear dynamical ideas into developmental biology in the form of the 'epigenetic landscape' - a general metaphor for the dynamics of the develop-mental process. The developmental paths of tissues and cells are seen to be constrained or canalized to 'flow' along certain valleys and not others due to the 'force' exerted on the landscape by the various gene products which define the fluid topography of the landscape. [7] This fluid topography contains multiple potential developmental pathways that may be realized as the result of 'fluctuations', or if the environmental conditions, the genes or gene products change. This metaphor has been made much more explicit recently by mathematician Peter Saunders (1992) who shows that the properties of the epigenetic landscape are 'common not just to developing organisms but to most nonlinear dynamical systems.'
A particular kind of nonlinearity which has made headlines recently is 'deterministic chaos': a complex dynamical behaviour that is locally unpredictable and irregular, which has been used to describe many living functions including the collective behaviour of ant colonies (see Goodwin, 1994). The unrepeatable patterns of brain activities that persuaded Freeman (1995) to declare brain science in crisis are typical of systems exhibiting deterministic chaos. Another putative example is the heart beat, which is found to be much more irregular in healthy people than in cardiac patients. [8] Physiologist Goldberger (1991) came to the conclusion that healthy heartbeat has 'a type of variability called chaos', and that loss of this 'complex variability' is associated with pathology and with aging. Similarly, the electrical activities of the functioning brain, apart from being unrepeatable from moment to moment, also contain many frequencies. But during epileptic fits, the spectrum is greatly impoverished (Kandel, Schwartz and Jessell, 1991). There is much current debate as to whether these complex variabilities associated with the healthy, functional state constitute chaos in the technical sense, so the question is by no means settled (Glass and Mackey, 1988).
A different understanding of the complex activity spectrum of the healthy state is that it is polychromatic (Ho, 1996c), approaching 'white' in the ideal, in which all the modes of energy storage are equally represented. It corresponds to the so-called f(l) = const. rule that Popp (1986) has generalized from the spectrum of light or 'biophotons' found to be emitted from all living systems. I have proposed that this polychromatic ideal distribution of stored energy is the state towards which all open systems capable of energy storage naturally evolve (Ho, 1994b). It is a state of both maximum and minimum in entropy content: maximum because energy becomes equally distributed over all the space-time modes (hence the 'white' ideal), and minimum because the modes are all coupled or linked together to give a coherent whole, in other words, to a single degree of freedom (Popp, 1986; Ho, 1993). In a system where there is no impedance to energy mobilization, all the modes are intercommunicating and hence all the frequencies will be represented. Instead, when coupling is imperfect, or when the subsystem, say, the heart, or the brain, is not communicating properly, it falls back on its own modes, leading to impoverishment of its activity spectrum. Living systems are necessarily a polychromatic whole, they are full of colour and variegated complexity that nevertheless cohere into a singular being.
One distinguishing feature of the living system is its exquisite sensitivity to weak signals. For example, the eye can detect single photons falling on the retina, and the presence of several molecules of pheromones in the air is sufficient to attract male insects to their appropriate mates. That extreme sensitivity of the organism applies to all levels and is the direct consequence of its energy self-sufficiency. No part of the system has to be pushed or pulled into action, nor be subjected to mechanical regulation and control. Instead, coordinated action of all the parts depends on rapid intercommunication throughout the system. The organism is a system of 'excitable media' (see Goodwin, 1994,1995), or excitable cells and tissues poised to respond specifically and disproportionately (i.e., nonlinearly) to weak signals because of the large amount of energy stored, which can thus amplify the weak signal into macroscopic action. It is by virtue of its energy self-sufficiency, therefore, that an organism is a sentient being - a system of sensitive parts all set to intercommunicate, to respond and to act appropriately as a whole to any contingency.
The organism is indeed free from mechanical determinism, but it does not thereby fall prey to indeterminacy. Far from surrendering its fate to the indeterminacy of nonlinear dynamics (or quantum theory, for that matter), the organism maximizes its opportunities inherent in the multiplicity of futures available to it. I have argued elsewhere that indeterminacy is really the problem of the ignorance of the external observer, and not experienced by the being itself, who has full knowledge of its own state, and can readily adjust, respond and act in the most appropriate manner (Ho, 1993). In a very real sense, the organism is free to decide its own fate because it is a sentient being who has moment to moment, up-to-date knowledge of its own internal milieu as well as the external environment.
This idea has become very concrete as the result of recent advances in biochemistry, cell biology and genetics. A molecular democracy of distributed control
There are thousands of enzymes catalyzing thousands of energy transactions and metabolic transformations in our body. The product of one enzyme is acted on by one or more other enzymes, resulting in a highly interconnected metabolic network. Henrik Kacser (1988) was among the first to realize that once we have a network, especially one as complicated as the metabolic network, it is unrealistic to think that there could be special enzymes controlling the flow of metabolites under all circumstances. He and a colleague pioneered metabolic control analysis, to discover how the network is actually regulated under different conditions.
After more than 20 years of investigation by many biochemists and cell biologists, it is now generally recognized that so-called 'control' is invariably distributed over many enzymes (and metabolites) in the network, and moreover, the distribution of control differs under different conditions. The metabolic network turns out to be a 'molecular democracy' of distributed control.
Recent studies have also revealed that energy mobilization in living systems is achieved by protein or enzyme molecules acting as 'flexible molecular energy machines' (see Ho, 1995a), which transfer energy directly from the point of release to the point of utilization, without thermalization or dissipation. These direct energy transfers are carried out in collective modes extending from the molecular to the macroscopic domain. The flow of metabolites is channeled coherently at the molecular level, from one enzyme to the next in sequence, in multi-enzyme complexes (see Welch and Clegg, 1987). At the same time, high voltage electron microscopy and other physical measurement techniques reveal that the cell is more like a 'solid state' than the 'bag of dissolved enzymes' that generations of biochemists had previously supposed (Clegg, 1984). Not only are almost all enzymes bound to an intricate 'microtrabecular lattice', but a large proportion of metabolites as well as water molecules are also structured on the enormous surfaces available. Aqueous channels are now thought to be involved in the active transport of solutes within the cell in the same way that the blood stream transport metabolites and chemical messengers within the organism (Wheatley and Clegg, 1991). Joseph Needham (1935) and his colleagues were already aware of all that some sixty years ago.
As Welch and Berry (1985) propose, the whole cell is linked up by 'long-range energy continua' of mechanical interactions, electric and eletrochemical fluxes and in particular, proton currents that form a 'protoneural network', whereby metabolism is regulated instantly and down to minute detail. In addition, the possibility that cells and tissues are also linked by electromagnetic phonons and photons is increasingly entertained (see Popp, Li and Gu, 1992; Ho, 1993; Ho, Popp and Warnke, 1994). As I shall show later, the cell (as well as organism) is not so much a 'solid state' as liquid crystalline. Living systems, therefore, possess just the conditions that favour the rapid propagation of influences in all directions, so that local and global can no longer be easily distinguished. Global phase transitions may often take place, which can be initiated at any point within the system or subsystem. Freeman and Barrie (1994) have described abrupt, phase-transition like changes that typically occur in the eeg of whole areas of the brain, recorded simultaneously with a large array of electrodes, for which no definite centre(s) of origin can be identified. [9]
Biology today remains dominated by the genetic paradigm. Genes are seen to be the repository of information that controls the development of the organism, but are otherwise insulated from the environment, and passed on unchanged to the next generation except for rare random mutations. The much publicized Human Genome Project is being promoted on that very basis. [10] Yet, the genetic paradigm has already been fatally undermined at least ten years ago, when a plethora of 'fluid genome' processes were first discovered, and many more have come to light since. These processes destabilize and alter genes and genomes in the course of development, some of the genetic changes are so well correlated with the environment that they are referred to as 'directed mutations'. Many of the genetic changes are then passed on to the next generation. I pointed out at the time that heredity can no longer be seen to reside solely in the DNA passed on from one generation to the next. Instead, the stability and repeatability of development - which we recognize as heredity - is distributed in the whole gamut of dynamic feedback interrelationships between organism and environment, from the socioecological to the genetic. All these may leave imprints that are passed on to subsequent generations, in the form of cultural traditions or artefacts, maternal or cytoplasmic effects, gene expression states, as well as genetic (DNA sequence) changes.
The organism is highly interconnected and intercommunicating at all levels extending from within the cell to the socioecological environment. It is on that account that the organism has freed itself from mechanistic controls of any kind. It is not a passive object at the mercy of random variation and natural selection, but an active participants in the evolutionary drama. [11] In constantly responding to and transforming its environment, it partakes in creating the possible futures of generations to come.
The concept of coherence has emerged within the past 20 years to describe the wholeness of the organism. The first detailed theory of coherence of the organism was presented by Herbert Fröhlich (1968; 1980) who argued that as organisms are made up of strongly dipolar molecules packed rather densely together (c.f. the 'solid state' cell), electric and elastic forces will constantly interact. Metabolic pumping will excite macromolecules such as proteins and nucleic acids as well as cellular membranes (which typically have an enormous electric field of some 10 [7]V/m across them). These will start to vibrate and eventually build up into collective modes, or coherent excitations, of both phonons and photons (sound and light) that extend over macroscopic distances within the organism and perhaps also outside the organism. The emission of electromagnetic radiation from coherent lattice vibrations in a solid-state semi-conductor has recently been experimentally demonstrated for the first time (Dekorsy et al, 1995). The possibility that organisms may use electromagnetic radiations to communicate between cells was already entertained by Soviet biologist Gurwitsch (1925) early this century.This hypothesis was revived by Popp and his coworkers in the late 1970s, and there is now a large and rapidly growing literature on 'biophotons' that are believed to be emitted from a coherent photon field (or energy storage field) within the living system (see Popp, Li and Gu, 1992).
We have indeed found that a single, one minute, exposure of synchronously developing early fruitfly embryos to white light results in the re-emission of relatively intense and prolonged flashes of light, some tens of minutes and even hours after the light exposure (Ho et al, 1992b). This is reminiscent of phase-correlated collective emission, or superradiance, in physical systems, although the timescale is orders of magnitude longer. For phase-correlation to build up over the entire population, one must assume that each embryo has a collective phase of all its activities, in other words, each embryo must be considered a highly coherent domain, despite its multiplicity of activities (Ho, Zhou and Haffegee, 1995). Actually, this is no different from the macroscopic phase correlations that are involved in the synchronous flashing of huge populations of fireflies (Strogatz and Mirollo, 1988), and in many physiological functions, such as limb coordination during locomotion (Collin and Stewart, 1992; Kelso, 1991) and coupling between heart rate and respiratory rate (Breithaupt, 1989). Under those conditions, whole limbs or entire circulatory and respiratory systems must be considered coherent domains which can maintain definite phase relationships with respect to one another.
During the same early period of development in Drosophila, exposure of the embryos to weak static magnetic fields also cause characteristic global transformation of the normal segmental body pattern to helical configurations in the larvae emerging 24 hours later (Ho et al, 1992a). As the energies involved are well below the thermal threshold, we conclude that there can be no effect unless the external field is acting on a coherent field where charges are moving in phase, or where magnetically sensitive liquid crystals are undergoing phase alignment globally (Ho, et al, 1994). Liquid crystals may indeed be the material basis of many, if not all aspects of biological organization (Ho et al, 1996).
Liquid crystals are phases of matter between the solid and the liquid states, hence the term, mesophases (DeGennes, 1974). Liquid crystalline mesophases possess long range orientational order (all the molecules pointing in the same direction), and often also varying degrees of translational order (the individual molecules keep to their positions to varying extents). In contrast to solid crystals, liquid crystals are mobile and flexible, and above all, highly responsive. They undergo rapid changes in orientation or phase transitions when exposed to electric or magnetic fields (Blinov, 1983) or to changes in temperature, pressure, pH, hydration, and concentrations of inorganic ions (Collings, 1990; Knight, 1993). These properties are ideal for organisms (Gray, 1993; Knight, 1993). Liquid crystals in organisms include all its major constituents; the lipids of cellular membranes, the DNA in chromosomes, all proteins, especially cytoskeletal proteins, muscle proteins, collagens and other macromolecules of connective tissues. These adopt a multiplicity of different mesophases that may be crucial for biological structure and function at all levels of organization (Ho et al, 1996) from channeling metabolites in the cell to pattern determination and the coordinated locomotion of whole organisms.
The importance of liquid crystals for living organization was recognized by Joseph Needham (1935) among others. He suggested that living systems actually are liquid crystals, and that many liquid crystalline mesophases may exist in the cell although they cannot then be detected. Indeed, there has been no direct evidence that extensive liquid crystalline mesophases exist in living organisms or in the cytoplasm until our recent discovery of a noninvasive optical technique (Ho and Lawrence, 1993; Ho and Saunders, 1994; Newton, Haffegee and Ho, 1995). This enables us to obtain high resolution and high contrast coloured images of live organisms based on visualizing just the kind of coherent liquid crystalline mesophases which Needham and others had predicted.
The technique effectively allows us to see the whole of the living organism at once from its macroscopic activities down to the phase alignment of the molecules that make up its tissues. Brilliant optical colours are generated which are specific for each tissue, dependent on the molecular structure and the degree of coherent alignment of all the molecules, even as the molecules are moving about busily transforming energy. This is possible because visible light vibrates much faster than the molecules can move, so the tissues will appear indistinguishable from static crystals to the light passing through so long as the movements of the constituent molecules are sufficiently coherent. With this imaging technique, one can see that the organism is thick with activities at all levels, which are coordinated in a continuum from the macroscopic to the molecular. And that is what the coherence of the organism entails.
These images also bring out another aspect of the wholeness of the organism: all organisms, from protozoa to vertebrates without exception, are polarized along the anteroposterior axis, so that all the colours in the different tissues of the body are at a maximum when the anteroposterior axis is appropriately aligned, and they change in concert as the organism is rotated from that position. The anteroposterior axis acts as the optical axis for the whole organism, which behaves in effect, as a single crystal. This leaves us in little doubt that the organism is a singular whole, despite the diverse multiplicity and polychromatic nature of its constituent parts.
The tissues not only maintain their crystalline order when they are actively transforming energy, the degree of order seems to depend on energy transformation, in that the more active and energetic the organism, the more intensely colorful it is, implying that the molecular motions are all the more coherent (Ho and Saunders, 1994; Ho et al, 1996). The coherence of the organism is therefore closely tied up with its energetic status, as argued in the beginning of this essay: the coherent whole is full of energy - it is a vibrant coherent whole.
The above considerations and observations show that the essence of organic wholeness is that it is distributed throughout its constituent parts so that local and global, part and whole are completely indistinguishable - the organism's activities being always fully coordinated in a continuum from the molecular to the macroscopic. That convinces me (as argued in detail in Ho, 1993, also Ho, 1996a) that there is something very special about the wholeness of organisms that is only fully captured by quantum coherence. [12] An intuitive appreciation of quantum coherence is to think of the 'I' that each and every one of us experience of our own being. We know that our body is a multiplicity of organs and tissues, composed of many billions of cells and astronomical numbers of molecules of many different kinds, all capable of working autonomously, and yet somehow cohering into the singular being of our private experience. That is just the stuff of quantum coherence. Quantum coherence does not mean that everybody or every element of the system must be doing the same thing all the time, it is more akin to a grand ballet, or better yet, a very large jazz band where everyone is doing his or her own thing while being perfectly in step and in tune with the whole.
A quantum coherent system maximizes both global cohesion and local freedom (Ho, 1993). This property is technically referred to as factorizability, the correlations between subsystems resolving neatly into self-correlations so that the subsystems behave as though they are independent of one another. It enables the body to be performing all sorts of different but coordinated functions simultaneously (Ho, 1995b). It also enables instantaneous, as well as noiseless intercommunication to take place throughout the system. [13] As I am writing, my digestive system is working independently, my metabolism busily transforming chemical energy in all my cells, putting some away in the longer term stores of fat and glycogen, while converting most of it into readily utilizable forms such as ATP. Similarly, my muscles are keeping in tone and allowing me to work the keyboard, while, hopefully, my neurons are firing in wonderfully coherent patterns in my brain. Nevertheless, if the telephone should ring in the middle of all this, I would turn to pick it up without hesitation.
The importance of factorizability is evoked by the movie character, Dr. Strangelove, portrayed by Peter Sellers as a megalomaniac scientist who wanted to rule the world. He was a wheelchair-bound paraplegiac, who could not speak without raising his arm in the manner of a Nazi salute. That is just the symptom of the loss of factorizability which is the hallmark of quantum coherence.
The coherent organism is, in the ideal, a quantum superposition of activities - organized according to their characteristic space-times - each itself coherent, so that it can couple coherently to the rest (Ho, 1995b; 1996a). This picture is fully consistent with the earlier proposal that the organism stores energy over all space-time domains each intercommunicating (or coupled) with the rest. Quantum superposition also enables the system to maximize its potential degrees of freedom so that the single degree of freedom required for coherent action can be instantaneously accessed.
The organism maximizes both local freedom and global intercommunication. One comes to the startling discovery that the coherent organism is in a very real sense completely free. Nothing is in control, and yet everything is in control. Thus, it is the failure to transcend the mechanistic framework that makes people persist in enquiring which parts are in control, or issuing instructions; or whether free will exists, and who choreographs the dance of molecules. Does 'consciousness' control matter or vice versa? These questions are meaningless when one understands what it is to be a coherent, organic whole. An organic whole is an entangled whole, where part and whole, global and local are so thoroughly implicated as to be indistinguishable, and each part is as much in control as it is sensitive and responsive. Choreographer and dancer are one and the same. The 'self' is a domain of coherent activities, in the ideal, a pure state that permeates the whole of our being with no definite localizations or boundaries, as Bergson has described.
The positing of 'self' as a domain of coherent activities implies the existence of an active whole agent who is free. I must stress that freedom does not entail the breakdown of causality as many commentators have mistakenly supposed. On the contrary, an acausal world would be one where it is impossible to be free, as nothing would be intelligible. Nevertheless, freedom does entail a new kind of organic causality that is nonlocal, and posited with the organism itself. It is the experience of perceptual feedback consequent on one's actions that is responsible for the intuition of causality (Freeman, 1990). However, it must not be supposed that the cause or consciousness is secreted from some definite location in the brain, it is distributed and delocalized throughout the system (c.f. Freeman, 1990).
Freedom in the present context means being true to 'self', in other words, being coherent. A free act is a coherent act. Of course not all acts are free, as one is seldom fully coherent. Yet the mere possiblity of being unfree affirms the opposite, that freedom is real,
'..we are free when our acts spring from our whole personality, when they express it, when they have that indefinable resemblance to it which one sometimes finds between the artist and his work.' [14]
The coherent 'self' is distributed and nonlocal - being implicated in a community of other entities with which one is entangled (Whitehead, 1925; see also Ho, 1993). Thus, being true to self does not imply acting against others. On the contrary, sustaining others sustains the self, so being true to others is also being true to self. It is only within a mechanistic Darwinian perspective that freedom becomes perverted into acts against others (see Ho, 1996e). The coherent 'self' can also couple coherently to the environment so that one becomes as much in control of the environment as one is responsive. The organism thereby partici-pates in creating its own possible futures as well as those of the entire community of organisms in the universe, much as Whitehead (1925) has envisaged.
I venture to suggest, therefore, that a truly free individual is a coherent being that lives life fully and spontaneously, without fragmentation or hesitation, who is at peace with herself and at ease with the universe as she participates in creating, from moment to moment, its possible futures.
An earlier draft of this paper was written for the occasion of the 6th Mind & Brain Conference, and I am grateful to Brian Goodwin and Peter Fenwick for making it happen. Afterwards, I felt so inspired by the discussions with the participants that I decided to write it up for publication. Thanks are also due to Geoffrey Sewell for stimulating discussions on coherence and bioenergetics and for keeping track of my physics; to Peter Saunders, Brian Goodwin, Michael Brown and Michael Clarke for their encouragement and support, and for drawing my attention to crucial publications and preprints. Invaluable suggestions for improving the manuscript came from the reviewers, Walter Freeman and Joseph Goguen.
1. The Theoretical Biology Club was an informal association of academics based in Cambridge University in the 1930s. Its membership was probably more extensive than I have indicated(see Mackay, 1994). Their project continued, to some extent, in a series of meetings organized by C.H. Waddington in the 1960s and 70s. The proceedings, published under the title,Towards a Theoretical Biology (Edinburgh University Press) were very influential among critics of mainstream neo-Darwinian theory of evolution, including myself. Four recent Waddington Memorial Conferences have been organized by Waddington's student, Brian Goodwin, and published as collected volumes (see Goodwin and Saunders, 1989; Stein and Varela, 1992). These helped to keep the project of the Theoretical Biology Club alive, and I count myself among the intellectual beneficiaries.
2. Cited in Ehrenber, 1967, p103.
3. Schrödinger, 1944, pp.70-71.
4. Schrödinger was criticized by both Pauling and Perutz over his non-rigorous use of 'negative entropy'. The exchanges are described by Gnaiger, 1994.
5. I explore the consequences of organic space-time for understanding some of the more paradoxical 'states of consciousness' in my book (Ho, 1993) and also in a forth-coming paper (Ho and Marcer, 1996).
6. The present conceptualization, based on thermodynamics, converges with the notion of autopoesis describing the living system as a unitary, self-producing entity, which Maturana and Varela (1987) derived from purely formal considerations.
7. Waddington's ideas in evolutionary theory is reviewed recently by Ho, 1996b.
8. This is comprehensively described by Goodwin (1995) in our Open University Third Level Course and accompanying video.
9. Elsewhere, it is argued that nonlocal intercommunication based on quantum coherence is involved in these simultaneous changes in brain activities (Ho and Marcer, 1996).
10. I have dealt with the socioeconomic implications as well as scientific issues of gene biotechnology and the Human Genome Project elsewhere Ho (1995c).
11. My colleagues and I have written against the reductionist tendencies of mainstream evolutionary theory since 1976, but see in particular, Ho and Saunders (1984); Pollard, J.W. (1984); Ho, M.W. (1986); Ho and Fox (1988). The issue of epigenetic, or Lamarckian inheritance has been thoroughly reviewed and documented recently by Jablonka and Lamb (1995). See also, Ho, M.W. (1996d).
12. Some aspects of brain activity can best be understood in terms of quantum coherence, independently of arguments given by Hameroff and Penrose (1995) who offer a specific mechanism for mediating coherence. The quantum coherence described in the present paper involves the whole system. When the system is coherent, nonlocal correlations can be established instantaneously, i.e., without delay. The largescale spatial coherence of brain activities observed by Freeman and Barrie (1994) may be indicative of such instantaneous intercom-munication. The relationship between quantum coherence, organic space-time and conscious experience is the subject of another paper (Ho and Marcer, 1966).
13. The coherent pure state (which is factorizable) is the prerequisite for instantaneous, lossless intercommunication, because the slightest change will give rise to a 'signal' passing between the uncorrelated factorizable parts. However, during intercommunication, factorizability is temporarily lost.
14. Bergson, 1916, p. 172.
Article first published 8/10/00
Comments are now closed for this article