Architectural Design Profile No. 129, New Science New Architecture (C. Jencks, ed.), pp. 44-51, Royal Academy of Art, 1997
I am told the comet in our sky visited us 4000 years ago. As it revolves once around the heaven, earth has revolved 4000 times around our sun, and human beings have gone from stone age to space age in 160 life cycles. The comet looks like a giant eye in the sky, now within our orbit and looking down on us, having seen, perhaps, many other worlds in far-flung reaches of the universe during its space odyssey. Do any of those other worlds contain beings that gaze back at it as we do? One begins to get a sense of a multitude of space-times entangled with our here and now. The here and now contains in its essence a myriad of there and thens. That is the real sense in which the "fullness of time" is to be understood. It is the reality of organic space-time that the mechanistic worldview has flattened out of existence.
Mechanical space and time are both linear, homogeneous, separate and local. In other words, both are infinitely divisible, and every bit of space or of time is the same as every other bit. A billiard ball here cannot affect another one there, unless someone pushes the one here to collide with the one there. Mechanical space-time also happens to be the space and time of the commonest "common-sensible" world in our mundane, everyday existence. It is the space-time of frozen instantaneity abstracted from the fullness of real process, rather like a still frame taken from a bad movie-film, which is itself a flat simulation of life. The passage of time is an accident, having no connection with the change in the configuration of solid matter located in space. Thus, space and time are merely coordinates for locating objects. One can go forwards or backwards in time to locate the precise objects at those particular points. In reality, we know that we can as much retrace our space-time to locate the person that was 30 or 50 years younger as we can undo the wrongs we have committed then. There is no simple location in space and time (Whitehead, 1925).
Psychoanalyst-artist Marian Milner (1957) describes her experience of "not being able to paint" as the fear of losing control, of no longer seeing the mechanical common-sensible separateness of things. It is really a fear of being alive, of entanglement and process in the organic reality that ever eludes mechanistic descripion. And yet, it is in overcoming the imposed illusion of the separateness of things that the artist/scientist enters into the realm of creativity and real understanding - which is the realm of organic space-time. Mechanical physics has banished organic space-time from our collective public consciousness, though it never ceases to flourish in the subterranean orphic universe of our collective unconscious and our subjective aesthetic experience. In a way, all developments in western science since Descartes and Newton may be seen as a struggle to reclaim our intuitive, indigenous notions of organic space-time, which, deep within our soul, we feel to be more consonant with authentic experience.
The mechanistic worldview indeed officially ended at the beginning of this century. Einstein's relativity theory broke up Newton's universe of absolute space and time into a multitude of space-time frames each tied to a particular observer, who therefore, not only has a different clock, but also a different map. Stranger still - for western science, that is, as it comes as little surprise to other knowledge systems, or to the artists in all cultures - quantum theory demanded that we stop seeing things as separate solid objects with definite (simple) locations in space and time. Instead, they are delocalized, indefinite, mutually entangled entities that evolve like organisms.
The profound implications of this decisive break with the intellectual tradition of previous centuries were recognized by a mere handful of visionaries. Among them, the French philosopher Henri Bergson (1916), and the English mathematician-philosopher Alfred North Whitehead (1925). Between them, they articulated an organicist philosophy in place of the mechanistic. Let me summarize some of what I see to be the major contrasts between the mechanical universe and the universe of organisms.
| Mechanical Universe | Organic Universe |
| Static, deterministic | Dynamic, evolving |
| Separate, absolute time, universal for observer (process)-dependent observers space-time frames | absolute space and Space-time inseparable, contingent all |
Inert objects with simple locations in space and time | Delocalized organisms with mutually entangled space-times |
| Linear, homogeneous space and time | Nonlinear, heterogeneous, multi- dimensional space-times |
| Local causation | Non-local causation |
| Given, nonparticipatory and hence, impotent observer | Creative, participatory; entanglement of observer and observed |
The contrasts are brought into sharper relief by considering the dif-ferences between mechanism and organism, or, more accurately, the opposition between a mechanical system and an organic system. First of all, a mechanical system is an object in space and time, whereas an organism is, in essence, of space-time. An organism creates its own space-times by its activities, so it has control over its space-time, which is not the same as external clock time. Secondly, a mechanical system has a stability that belongs to a closed equilibrium, depending on controllers, buffers and buttresses to return the system to set, or fixed points. It works like a non-democratic institution, by a hierarchy of control: a boss who sits in his office doing nothing (bosses are still predominantly male) except giving out orders to line managers, who in turn coerce the workers to do whatever needs to be done. An organism, by contrast, has a dynamic stability, which is attained in open systems far away from equilibrium. It has no bosses, no controllers and no set points. It is radically democratic, everyone participates in making decisions and in working by intercommunication and mutual responsiveness. Finally, a mechanical system is built of isolatable parts, each external and independent of all the others. An organism, however, is an irreducible whole, where part and whole, global and local are mutually implicated.
I hope you are sufficiently persuaded that we need a radically new way of understanding the organism, if not the whole of nature, as Whitehead intimates. In this project, we - each and everyone of us - are especially privileged, because we are ourselves organisms and know in intimate, exquisite detail, what it is to be alive.
The vast majority of scientists as well as the general public have remained untouched by this conceptual revolution. Quantum theory itself sits uneasily and paradoxically between the necessary limits of a mechanical description (in quantum mechanics) and the elusive, organic reality that remains ever out of reach. Mathematics and physics have recently broken out of the strict mechanistic mould to explore the 'organic' realm (see Ho, 1993; 1996a, also Saunders, this volume). In mathematics, computations have made accessible previously intractable problems in nonlinear dynamics, fractal geometry and chaos. In the mean time, physics has witnessed an astonishing inventory of empirical successes - high temperature superconductivity, quantum coherence and nonlocal quantum superposition of states - even as theoretical descriptions have lagged far behind. It is precisely at the point where theoretical description fails to capture the organic freedom of reality that contemporary science is at its most captivating. It is the realm of imagery where scientist and artist meet, and where no one who is not both can enter.
The end of mechanistic biology
Mainstream biology is left far behind. It is clinging fast to the mechanistic era. The discovery of the DNA double-helix in the late 1950s, which has made its permanent mark on the public consciousness, was the climax to a century of mechanistic, reductionist biology - the idea that the whole is the sum of its parts, that cause and effect are simply related, and can be neatly isolated. The discovery ended the quest for the material basis of the units of heredity - the genes - that are supposed to determine the characters of organisms and their offspring, thus firmly establishing the predominance of the genetic determinist paradigm. The subsequent flowering of molecular biology gave rise to the present era of recombinant DNA research and commercial genetic engineering biotechnology.
What few people realize is that the very successes of recombinant DNA research have completely undermined the foundations of the genetic determinist paradigm, at least ten years ago. There has indeed been a revolution in genetics which exactly parallels the transition between mechanical and quantum physics. The new genetics signals the final demise of mechanistic biology, and is consonant with the diametrically opposite, organicist perspective which has been emerging in the rest of science. The contrast between the old, pre-recombinant DNA genetics and the new genetics is presented below.
| The Old Genetics | The New Genetics |
| Genes determine characters in a linear, additive way | Genes function in a complex, nonlinear, multidimensional network - the action of each gene ultimately linked to that of every other. |
| Genes and genomes are stable and except for rare random fluid, mutations, are passed on un- changed to the next generation | Genes and genomes are dynamic and they can change in the course of development, and as the result of feedback metabolic regulation |
| Genes and genomes cannot be changed directly in response to the environment | Genes and genomes can change directly in response to the environment, these changes being inherited in subsequent generations |
| Genes are passed on vertically i.e., as the result of inter- breeding within the species, each species constituting an isolated 'gene pool' | Genes can also be exchanged horizontally between individuals from the same or different species |
The parallel to the transition from classical to quantum physics is best illustrated by focussing on the concept of the "gene" (see Ho, 1997a for details). In the old genetics, the "gene" is a continuous stretch of DNA, with a particular base sequence, and a constant, simple location in the genome, that specifies, via a non-overlapping triplet code, the amino-acid sequence of a single protein. The amino-acid sequence of the protein, in turn, determines its function in the organism. The genetic code is universal, and there is a "one-way information flow" from DNA to an intermediary "messenger" RNA to the protein, and no reverse information flow is possible. This was the notion of a definite, isolatable gene, specifying a function independently of the cellular and environmental context.
The cracks in the old edifice first appeared when reverse information flow was found to occur from RNA back to DNA. Then, the genetic code was discovered to be overlapping and non-universal. Next came a succession of revelations showing that the gene itself has no well-defined continuity nor boundaries, the expression of each gene being ultimately dependent on, and entangled with every other gene in the genome. Far from the one-way information flow that is supposed to proceed from DNA to RNA to protein and on to the rest of the organism, gene expression is subject to influences and instructions from the cellular and environmental contexts. The gene can be recoded, or edited by the cell, it can get silenced, or converted to a different sequence. Genome organization is infinitely variable, dynamic and fluid. Genes mutate frequently, small and large rearrangements take place, genes jump around, sequences are added or deleted, they get amplified thousands and hundreds of thousands of times or they get contracted. These changes may take place as part of normal development or they occur repeatably in response to environmental challenges. Some of the genetic changes are so specific that they are referred to as "directed mutations" or "adaptive mutations". Genes can even jump horizontally, by infection, between species that do not interbreed. Genes and genomes are in reality, dynamic, delocalized, mutually entangled and part of larger wholes. In short, biology has been catapulted, over the heads of the old guard, into the new age of the organism.
I have given a good indication of what the new "physics of the organism" might look like in an earlier book (Ho, 1993) and in other recent publications (Ho, 1995,a,b; 1996a,b). In the rest of this paper, I shall outline a theory of the organism, ending with a few remarks on certain key aspects that are most relevant to organic, as opposed to mechanistic forms: organic stability, organic space-time and the integral delocalization of organic forms.
There are 75 trillion cells in our body, made up of astronomical numbers of molecules of many different kinds. How can this huge conglomerate of disparate cells and molecules function so perfectly as a coherent whole? How can we summon energy at will to do whatever we want? And most of all, how is it possible for there to be a singular "I" that we all feel ourselves to be amid this diverse multiplicity and plurality?
To give you an idea of the coordination of activities involved, imagine an immensely huge superorchestra playing with instruments spanning an incredible spectrum of sizes from a piccolo of 10-9 metre up to a bassoon or a bass viol of 1 metre or more, and a musical range of seventy-two octaves. The amazing thing about this superorchestra is that it never ceases to play out our individual songlines, with a certain recurring rhythm and beat, but in endless variations that never repeat exactly. Always, there is something new, something made up as it goes along. It can change key, change tempo, change tune perfectly, as it feels like it, or as the situation demands, spontaneously and without hesitation. Furthermore, each and every player, however small, can enjoy maximum freedom of expression, improvising from moment to moment, while maintaining in step and in tune with the whole.
I have just given you a theory of the quantum coherence that underlies the radical wholeness of the organism. It is a special wholeness that involves total participation, and maximizes both local freedom and global cohesion. It involves the mutual implication of global and local, of part and whole, from moment to moment. It is on that basis that we can have a sense of ourselves as a singular being, despite the diverse multiplicity of parts. That is also how we can perceive the unity of the here and now, in an act of "prehensive unification"(Whitehead, 1925). Artists like scientists, depend on the same exquisite sense of prehensive unification, to see patterns that connect apparently disparate phenomena.
In order to add corroborative details to my story, however, I shall give a more scientific narrative involving some easy lessons in thermodynamics and quantum theory. It begins with energy relationships.
Textbooks tell us that living systems are open systems dependent on energy flow. Energy flows in together with materials, and waste products are exported as well as the spent energy that goes to make up entropy. And that is how living systems can, in principle, escape from the second law of thermodynamics. The second law, as you may know, encapsulates the fact that all physical systems run down, ultimately decaying to homogeneous disorganization when all useful energy is spent, or converted into entropy. But how do living systems manage their antientropic existence?
I have suggested (Ho, 1996a,b) that the key to understanding how the organism overcomes the immediate constraints of thermodynamics is in its capacity to store the incoming energy, and in somehow closing the energy loop within to give a reproducing, regenerating life cycle (see Figure 1). The
energy, in effect, goes into complex cascades of coupled cyclic processes within the system before it is allowed to dissipate to the outside. These cascades of cycles span the entire gamut of space-times from slow to fast, from local to global, that all together, constitutes the life-cycle (see Figure 2 for an intuitive picture). Each cycle is a domain of coherent energy storage - coherent energy is simply energy that can do work because it is all coming and going together, as opposed to incoherent energy which goes in all directions at once and cancel out, and is therefore, quite unable to do work.
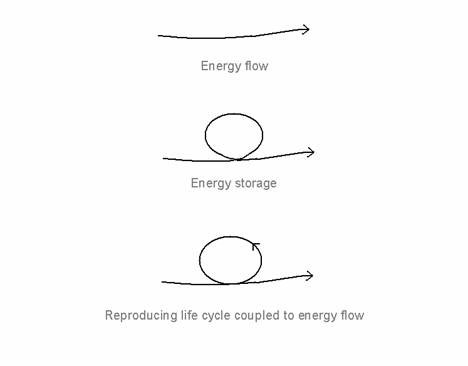
Figure 1. Energy flow, energy storage and the reproducing life-cycle
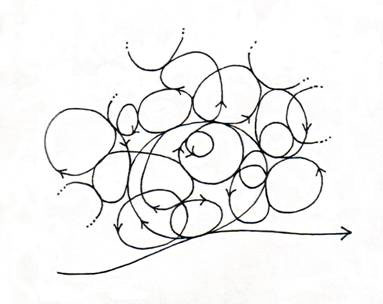
Figure 2. The many-fold cycles of life coupled to energy flow
Coupling between the cycles ensures that the energy is transferred directly from where it is captured or produced, to where it is used. In thermodynamic language, those activities going thermodynamically down-hill, and therefore yielding energy, are coupled to those that require energy and go thermodynamically uphill. This coupling also ensures that positive entropy generated in some space-time elements is compensated by negative entropy in other space-time elements. There is, in effect, an internal energy conservation as well as an internal entropy compensation. The whole system works by reciprocity, a cooperative give and take which balances out over the system as a whole, and within a sufficiently long time. The result is that there is always coherent energy available in the system. Energy can be readily shared throughout the system, from local to global and vice versa, from global to local, which is why, in principle, we can have energy at will, whenever and wherever it is needed. The organism has succeeded in gathering all the necessary vital processes into a unity of coupled non-dissipative cycles spanning the entire gamut of space-times up to and including the life-cycle itself, which effectively feeds off the dissipative irreversible energy flow (see Figure 3).
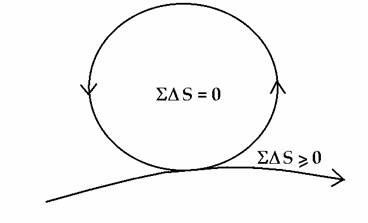
Figure 3. The organism frees itself from the contraints of energy conservation and the second law of thermodynamics
But how can energy mobilization be so perfectly coordinated? That is a direct consequence of the energy stored, which makes the whole system excitable, or highly sensitive to specific weak signals. It does not have to be pushed and dragged into action like a mechanical system. Weak signals originating anywhere within or outside the system will propagate throughout the system and become automatically amplified by the local energy stored, often into macroscopic action. Intercommunication can proceed very rapidly, especially because organisms are completely liquid crystalline. Let me explain.
Several years ago, we discovered an optical technique that enables us to see living organisms in brilliant interference colours generated by the liquid crystallinity of their internal anatomy. We found that all live organisms are completely liquid crystalline - in their cells as well as the extracellular matrix, or connective tissues (see Ho et al, 1996; Ross et al, 1997). Liquid crystals are states of matter between solid crystals and liquids. Like solid crystals, they possess long-range orientation order, and often, also varying degrees of translational order (or order of motion). In contrast to solid crystals, however, they are mobile and flexible and highly responsive. They undergo rapid changes in orientation or phase transitions when exposed to weak electric (or magnetic) fields, to subtle changes in pressure, temperature, hydration, acidity or pH, concentrations of inorganic molecules or other small molecules. These properties happen to be ideal for making organisms, as they provide for the rapid intercommunication required for the organism to function as a coherent whole. Some images of live organisms taken from video-recordings are shown in Figure 4.
Figure 4. The liquid crystalline organism. Images are still frames from a video-recording of live organisms viewed with a special polarized light microscopy technique which detects liquid crystalline regimes, a,b, successive frames of a first instar fruitfly larva about to hatch; c,d successive frames of the first instar fruitfly larva shortly after hatching; e,f successive frames of the trunk region of the brine shrimp.
What you are seeing is the whole of the organism at once, from its macroscopic activities down to the long-range order of the molecules that make up its tissues. The interference colours generated depend on the structure of the particular molecules, which differ for each tissue, and their degree of coherent order. The principle is exactly the same as that used in detecting mineral crystals in geology. But, with the important difference that the living liquid crystals are dynamic through and through, as the molecules are all moving about busily transforming energy and material in the meantime. So, how can they still appear crystalline?
Because visible light vibrates much faster than the molecules can move, the tissues will appear indistinguishable from static crystals to the light transmitted, so long as the movements of the constituent molecules are sufficiently coherent. Actually, the most actively moving parts of the organism are always the brightest, implying that their molecules are moving all the more coherently. With our optical technique, therefore, one can see that the organism is thick with coherent activities at all levels, which are coordinated in a continuum from the macroscopic to the molecular. That is the essence of the organic whole, where local and global, part and whole are mutually implicated at any time and for all times.
These images draw attention to the wholeness of the organism in another respect. All organisms - from protozoa to vertebrates without exception - are polarized along the anterior-posterior axis, or the oral-adoral axis, such that all the colours in the different tissues of the body are at a maximum when the axis is appropriately aligned in the optical system, and they change in concert as the axis is rotated from that position. The fruitfly larva has cleverly demonstrated that for us by curling its body around in a circle (Figure 4 c,d).
As said before, intercommunication can proceed very rapidly through the liquid crystalline continuum of cells and connective tissues that make up the organism. In the limit of the coherence time and coherence volume of energy storage - the time and volume respectively over which the energy remains coherent - intercommunication is instantaneous or nonlocal. There is no time-separation within the coherence volume, just as there is no space-separation within the coherence time. Because the organism stores coherent energy over all space-times, it has a full range of coherent space-times, which are furthermore, all coupled together. Thus, there is a possibility for nonlocal intercommunication throughout the system. In the ideal, the system is a quantum superposition of coherent activities, constituting a "pure coherent state" that maximizes both local freedom and global cohesion, in acordance with the factorizability of the quantum coherent state (Ho, 1993, 1996a,b). Factorizability means that the different parts are so perfectly intercorrelated that the intercorrelations resolve neatly into products of the self-correlations. So the parts behave as though they are independent of one another. This is the radical nature of the organic whole (as opposed to the mechanical whole), where global cohesion and local freedom are both maximized, and each part is as much in control as it is sensitive and responsive.
The "whole" is thus a domain of coherent activities, constituting an autonomous, free entity (see Ho, 1996a), not because it is separate and isolated from its environment, but precisely by virtue of its unique entanglement of other organic space-times in its environment. In this way, one can see that organic wholes are nested as well as entangled individualities. Each can be part of a larger whole, depending on the extent over which coherence can be established. So, when many individuals in a society have a certain rapport with one another, they may constitute a coherent whole, and ideas and feelings can indeed spread like wildfire within that community. In the same way, an ecological community, and by extension, the global ecology may also be envisaged as a super-organism within which coherence can be established in ecological relationships over global, geological space-times (see Ho, 1993, 1997b).
The ideal quantum coherent state involving the whole system is a global attractor to which the system tends to return when it is perturbed, but as the system is always open, it will invariably be taken away from the totally coherent state. So here is how space-time, as well as entropy or time's "arrow", is generated (see Ho, 1993). It is generated in proportion to the incoherence of actions taken. The more the actions taken are at odds with the coherence of the system, the more time, and entropy, is generated, and the more the system ages. Thus, the biological age of an organism may literally be quite different from the age as measured by external clock-time. In the same way, the earth itself can be aging much faster on account of our incoherent actions within it. On the other hand, we may indeed enter a state of delocalized timelessness when we achieve a high degree of coherence. Some of us get an inkling of that during an aesthetic experience, or alternatively, a religious experience.
Several people have asked me whether it is possible to get younger. My first reaction was no, because for all real processes, according to the textbook, entropy is greater than or equal to zero. On further reflection, however, I think the answer has to be yes. It follows from the principle of internal entropy compensation in an organic system, where negative as well as positive entropy can be generated, and also because past and present, as well as present and future, can be nonlocally interconnected. The challenge is indeed to set ourselves and the earth back on a possibly rejuvenating, or at any rate, anti-entropic and self-sustaining course (see Ho, 1997b).
Organic space-time is tied to activity, and as elaborated above, these activities are fundamentally anti-entropic on account of their tendencies towards coherence. The organism is thus a coherent space-time structure engendering nonlocal interconnectedness. What is the nature of this structure?
There are several lines of recent evidence converging to a new picture of the "texture of reality" (see Stewart, 1989) suggesting that organic space-time does have a structure, and that this structure is fractal. One of the most exciting discoveries in recent years, which has given rise to the "science of complexity" is that natural processes and natural structures have fractal dimensions. That means they have dimensions in between the one, two or three to which we are accustomed. Fractals capture a new kind of order characterized by self-similarity - the similarity of part to whole over many different scales. Snowflakes, clouds, ferns, coastlines branching patterns of blood vessels, and the "cytoskeleton" inside each cell are all examples of fractal structures. Natural processes, from weather patterns to the healthy heart-beat and electrical activities of the brain, similarly, exhibit "chaotic dynamics" that when spatialized as a "Poincaré section" (see Stewart, 1989), gives rise to "strange attractors" that again have fractal dimensions. If space-time is indeed generated by processes as I have proposed here, then it should also exhibit fractal dimensions, or more accurately, multi-fractal dimensions. This is the basis of the "space-time differentiation" of organisms (see Ho, 1993).
According to Nottale (1996) and others, the whole of present day physics relies on the unjustified assumption of the differentiability of the space-time continuum, which stems from the classical domain, whereas Feynman and Hibbs (1965) have already shown that the typical path of a quantum particle is continuous, but nondifferentiable. This is the failure of present-day physical description to capture the organic quantum reality that I have alluded to earlier. For the description is still based on a mathematical representation of space-time as continuous and homogeneous, i.e., as infinitely divisible or "differentiable". It so happens that a structure that satisfies the requirement for continuity and non-differentiability is also fractal. Nottale (1996) writes,
"Giving up the hypothesis of differentiability has an important physical consequence: one can show that curves, surfaces, volumes and, more generally, spaces of topological dimension DT, which are continuous but non-differentiable, are characterized by a length, an area and, more generally a DT measure which becomes explicitly dependent on the resolution at which they are considered, ... and tends to infinity when the resolution interval e tends to zero. In other words, a non-differentiable space-time continuum is necessarily fractal....This result naturally leads to the proposal of a geometric tool adapted to construct a theory based on such premises, namely, fractal space-time."
The author then proceeds to describe a new approach that generalizes Einstein's principle of relativity to scale transformations. Specifically, the equations of physics are required to keep their form under scale transformation, i.e., to be scale covariant. It allows physicists to recover quantum mechanics as mechanics on a fractal space-time, in which Schrödinger's wave equation is a geodesic equation.
I wonder if that is not the beginning of an approach towards the quantization of space-time which, I believe, is a necessary consequence of the quantization of action that Planck's constant already involves. This quantized space-time is also Bergson's "duration", which expresses the indivisibile multiplicity of our subjective experience of organic process (see Ho, 1993). It is the experience of processes cascading through the continuous scales of fractal space-times that are all coupled together or entangled through the coherence of the "ground" or asymptotic state, over which the scale covariance is defined.
Organic architecture is nothing new. As Jencks (1995) points out, "'Organic unity, where not a part can be added or subtracted except for the worse' are injunctions that have rebounded through the halls of building sites for 2000 years." The artists have been well ahead of scientists after all. Jencks touches on some of the themes developed in this Chapter in his grand panoramic sweep of how the new "science of complexity" is changing architecture and culture. However, as with quantum theory itself, much of the science of complexity is still mechanism aspiring towards organism. So, perhaps there is an excuse for me to give in to the temptation of trying to imagine what organic architecture would be like, based on the new view of the organism just presented, and to make connections with some well-known and lesser known concepts in established organic architecture.
One question which arose (for some of us) in the wake of the discoveries of the new genetics is, how do organisms and species maintain their stability when genes and genomes are so mutable and fluid? That is a question on the nature of organic stability in general.
The conventional, neo-Darwinian explanation is that natural selection is always at work to select out those that are unstable, and hence "unfit", so only those that are sufficiently stable remain to propagate offspring like themselves. A neo-Darwinian account of architecture, might similarly explain that buildings are selected for stability - those that were not stable simply fell down and eliminated themselves, leaving the stable ones for us to admire and to imitate.
I do not know how that explanation fares in architecture, but it certainly fails to account for the responsiveness of organisms, including their genes and genomes, to environmental and physiological changes (see Ho, 1997a). The stability of organisms and species is dependent on the entire gamut of dynamic feedback interrelationships extending from the socio-ecological environment to the genes. Genes and genomes must also adjust and respond, and if necessary, to change, in order to maintain the stability of the whole. As said above, the stability of organisms is diametrically opposite to the stability of mechanical systems. Mechanical stability - which includes that of so-called "cybernetic" systems - belongs to a closed, static equilibrium, maintained by the action of controllers, buffers or buttresses designed to return the system to set points. Organic stability, on the other hand, is a dynamic balance attained in open systems far away from equilibrium, without controllers or set points, but by means of intercommunication and mutual responsiveness. The stability of organisms depends on all parts of the system being informed, participating and acting appropriately in order to maintain the whole.
Organic stability is therefore delocalized throughout the system, via symmetrically commuting parts, each of which changes in response to all the others and to the environment. I am reminded of Cecil Balmond's constructions (this volume), his "free forms" which defy gravity. Organic stability is in the dynamic integrity of the whole. I can imagine the stresses and strains distributing and ever-shifting from one part to another in cycles of correlated reciprocity. If these forms were made of transparent, liquid crystalline material, as living organisms are, one might see a beautiful display of ever-changing colour patterns reflecting the shifting patterns of stresses and strains, as the structure communicates with its environment, just as one can see in real organisms.
Organic forms are supported and sustained by their relationship to the environment. Liquid crystals, in particular, are constantly evolving embodiments of their changing environments, their surfaces are invariably curved and flexible, hence the study of their structure is referred to as "flexi-crystallography" by crystallographer Alan MacKay. Liquid crystals go through many abrupt phase transitions, each "phase" being itself a continuum of more subtle variations. The phases are all minimum energy surfaces separating an "inside" from the "outside", though the inside and the outside can be so thoroughly interdigitated that it becomes a major problem in topology to disentangle them. The infinite variety of intricately sculpted exoskeletons of radiolarians are mineral deposits templated by different liquid crystalline formations. Liquid crystalline structures have already inspired certain architectural designs, such as the carpark in the National University of Mexico (Alan Mackay, personal communication).
Whereas a mechanical form is located in space and persists (or not) in time, an organic form, by contrast, is a space-time structure; to be exact, a coherent space-time structure. An organic form creates space-time, increasing its space-time differentiation in the course of development and in evolution. Being in an organic form is to partake of its distinctive space-time, and its possibility for nonlocal interconnections over multiple dimensions. Jencks (1995) points out that virtually all those who referred to 'organic architecture', including classicists such as Vitruvius and Alberti, and modernists, such as Gropious and Wright, insisted on work that shows fractal self-similarity, or 'unity with variety'.
I have proposed above that organic space-time is fractal because it arises out of natural processes which are fractal. Fractal architecture, therefore, are unique creations of organic space-times that extend and enhance our experience as organisms. I am captivated by Bruce Goff's plan of his Bavinger House (see Jencks, 1995, p.45). It exhibits the dynamic, nonlocal inter-connections and the multiple resonances of the fractal, organic whole, simultaneously unfolding and enfolding, diverging and converging in the gesture of life itself. I imagine sounds taking on added dimensions of musicality and coherence within this structure.
Another aspect of organic space-time is its complexity, or space-time differentiation (see Ho, 1993). This corresponds, I think, to Jencks' (1995) concept of the "organizational depth" of an architecture - the "density with which things are linked" - which counters the 'depthless present' of modernist architecture by "building in time". The depth of organic space-time is not just a nestedness but a special kind of superpositionand entanglement. A fine example of space-time entanglement is Rem Koolhaas' library in Jussieu University, Paris (see Jencks, 1995, p. 87). It is a continuous linear route traversing a stack of near-horizontal planes connecting one level to the next; the whole floor is one unbroken multi-level ramp through which weaves "a grid of columns and randomized incidents".
Of course, organic architecture is not restricted to fractal constructions, just as organic processes can undergo global phase transitions or catastrophic changes. In terms of space-time structure, phase transitions would correspond to major reorganizations of the system, giving rise to a new Schrödinger's wave equation, or new "geodesic" (see above). Jencks has explored nonlinear and catastrophic forms to much effect in his interior and exterior designs and in landscaping.
Finally, it must be stressed that an organism is an unique embodiment of its environment, that arises out of an uninterrupted act of "prehensive unification" (Whitehead, 1925), a Bergsonian duration. Put in another way, the organism is an unique, integral space-time entangling a multitude of space-times. It simultaneously creates its own space-time while being constitutive of other space-times. As a work of art, the organic architecture is more than an icon or a symbol. It is a coherent superposition and entanglement that gives nonlocal access to the diverse multiplicity of space and times that constitute its integral whole. That may be the real challenge to organic architecture.
This essay benefited from stimulating discussions with Charles Jencks and Cecil Balmond, and with Alan Mackay. Philipe Herbomel drew my attention to Nottale's papers on fractal space-time by kindly sending the xerox copies. Part of this article was first presented as a public lecture "A theory of the organism and organic space-time" at a Conference on Time and Timelessness, Dartington Hall, April 9-13, 1997. I was much inspired by the occasion, by the responses of the audience, and by composer and scholar, Edward Cowie, who introduced my lecture. Julian Haffegee exercised great skill in preparing the colour images of Figure 4.
Article first published 26/07/00
Comments are now closed for this article